






حركية النيتروجين في التربة
مقدمة:
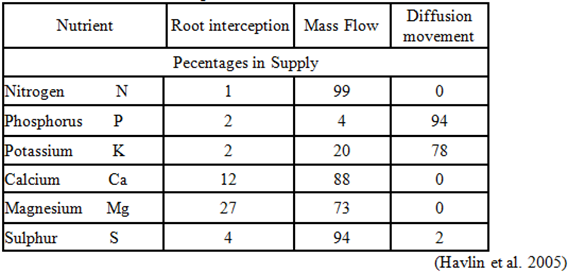
تعتمد النباتات في حصولها على العناصر ضعيفة الحركة مثل الفوسفور المعدني على ما يسمى بالاعتراض المباشر للجذور. إلا أن الحال يختلف بالنسبة للعناصر سريعة الحركة مثل النترات فإن معظم الكمية الممتصة تصل إلى الجذور محمولة مع جريان الكتلة والإنتشار، حيث أن الكمية الممتصة بواسطة الإعتراض المباشر من قبل الجذور لا تتجاوز 1% من إجمالي الكمية الممتصة وبالمقابل فإن جريان الكتلة يتعلق بعملية النتح وذلك لخلق القوة الدافعة لمحلول التربة ليتحرك نحو المجموع الجذري.
في المقابل عندما ينخفض محتوى التربة من الرطوبة فإن الدور الأبرز في تلبية احتياجات النبات من النيتروجين يعود لعملية الانتشار والذي يتعلق بعدة عوامل منها التعرج في تركيز الأيون ومعامل انتشاره في محلول التربة، ويتعلق بتأثير حجم الأيون وشحنته ولزوجة الماء ودرجة الحرارة ودرجة تعرج المسار والسعة التنظيمية للتربة. وبالتالي فإن معامل انتشار الأمونيوم أقل بحوالي 10 - 100 مرة من معامل انتشار النترات (Owen and Jones, 2001).
وبذلك تكون حركية النترات في محلول التربة أعلى من حركية الأمونيوم، ومما يجعل الأمونيوم أقل عرضة للفقد من آفاق التربة بعملية الإنغسال مدمصاً على غرويات التربة، ولكن ذلك يجعله أقل إتاحة للنبات مقارنة بالنترات (Colmer and Bloom, 1998).
يوضح الجدول التالي آليات انتقال العناصر إلى جذور النبات كنسبة مئوية من الكمية الكلية التي يمتصها النبات، والمتلخصة بالإعتراض الجذري Root interception والتدفق الكتلي Mass Flow إضافة إلى آلية الإنتشار في محلول التربة Diffusion movement.
امتصاص النيتروجين:
معظم الأنواع النباتية قادرة على امتصاص وتمثيل كلا الشكلين الآزوتيين (النترات والأمونيوم)، ولكن درجة تفضيل أحد الأشكال الآزوتية على الأخر يعتمد على صفات النوع النباتي، ودرجة تكيفه في الوسط المحيط، أو ظروف التربة. ففي الترب الرطبة وذات الرقم الهيدروجيني المنخفض نسبياً، فإن الشكل الأمونياكي هو السائد وبالتالي فإن النباتات التي تنمو في هذه الظروف تكيفت لامتصاص الأمونيوم. بينما النباتات التي تنمو في الترب ذات الرقم الهيدروجيني المائل للقلوية وذات تهوية جيدة فإن الشكل النتراتي يعتبر السائد والمفضل للنباتات التي تنمو في هذه الظروف، ولكن بشكل عام فإن معظم المحاصيل تفضل تواجد كلا الشكلين معاً (Daubresse et al., 2010).
الشكل الآتي يوضح الإمتصاص النشط لشوارد النترات والأمونيوم بعكس التدرج الكهروكيميائي والتي يحتاج عملها إلى طاقة، حيث نلاحظ ازدياد محتوى الجذور من الكربوهيدرات التي هي مصدر الطاقة لامتصاص العناصر الغذائية من خلال عملية التنفس، حيث يقوم ATPase المرتبط بالغشاء الخلوي بتفكيك الأدينوزين ثلاثي الفوسفات وإعطاء طاقة وأيون الفوسفويل الموجب.
![]()
ولا يلبث هذا الأيون الموجب غير المستقر أن يتفاعل مع الماء لإعطاء بروتون وفق المعادلة:
![]()
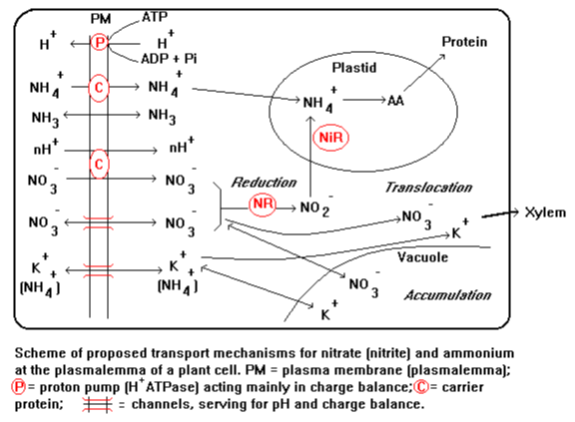
الشكل 1: يوضح تفاعلات الامتصاص النشط الفعال على سطوح جذور النبات الملامسة للتربة
حيث تقوم المضخة البروتونية المرتبطة بالأغشية البلازمية بضخ البروتونات من السايتوبلازم إلى المحلول الخارجي وتتشكل بالتالي قوة دافعة لنقل الكاتيونات والأنيونات عبر الأغشية البلازمية إلى داخل الجذور. ونلاحظ عند تدفق البروتونات إلى الخارج حصول تدرج في الأغشية البلازمية نحو الداخل، حيث يبقى الأيون السالب الممثل بشوارد الهيدروكسيل في السايتوبلازم، مما يؤدي لنشوء جهد كهربائي سالب للخلية مقارنة بالوسط الخارجي مما يؤدي إلى جذب شوارد الأمونيوم
قم بتنزيل تطبيق eMufeed Android الآن
